骨格筋の起源 |
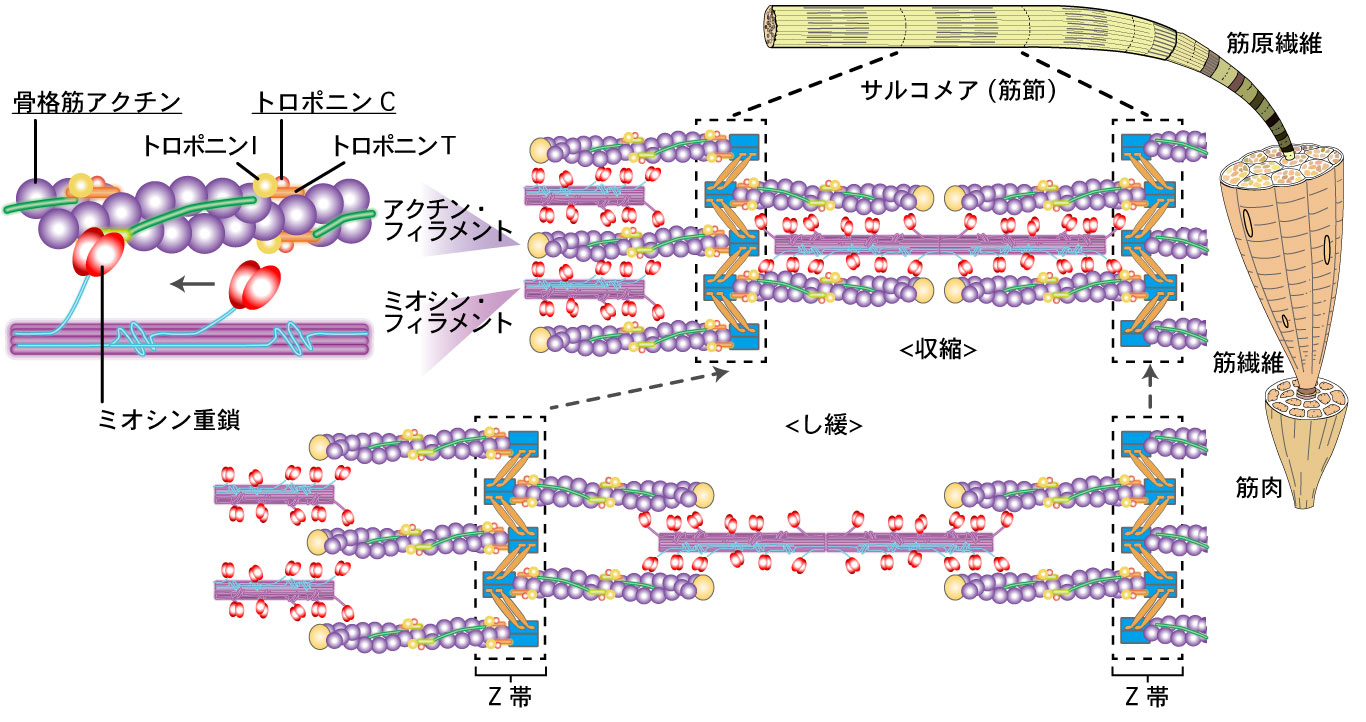 |
| 図1. 骨格筋の構造と滑り説. |
そこで我々は,骨格筋に相当する筋肉のない水腔動物のゲノムにも骨格筋遺伝子と同じ遺伝子が存在する可能性を考え,水腔動物を含めた後口動物全系統のゲノムデータから骨格筋タンパク質の遺伝子を探した.すると,由来の明らかになった 16 タンパク質のうち (図2右),速い動きに特化した骨格筋アクチンとトロポニン C の遺伝子が,水腔動物には存在しなかった.アミノ酸配列を比較したところ,骨格筋アクチンは脊索動物の祖先で新たに得られたものであった.一方トロポニン C は,脊索動物だけでなく前口動物も持っているが,水腔動物にはないことがはっきりしたので,水腔動物の祖先で失われたことになる.
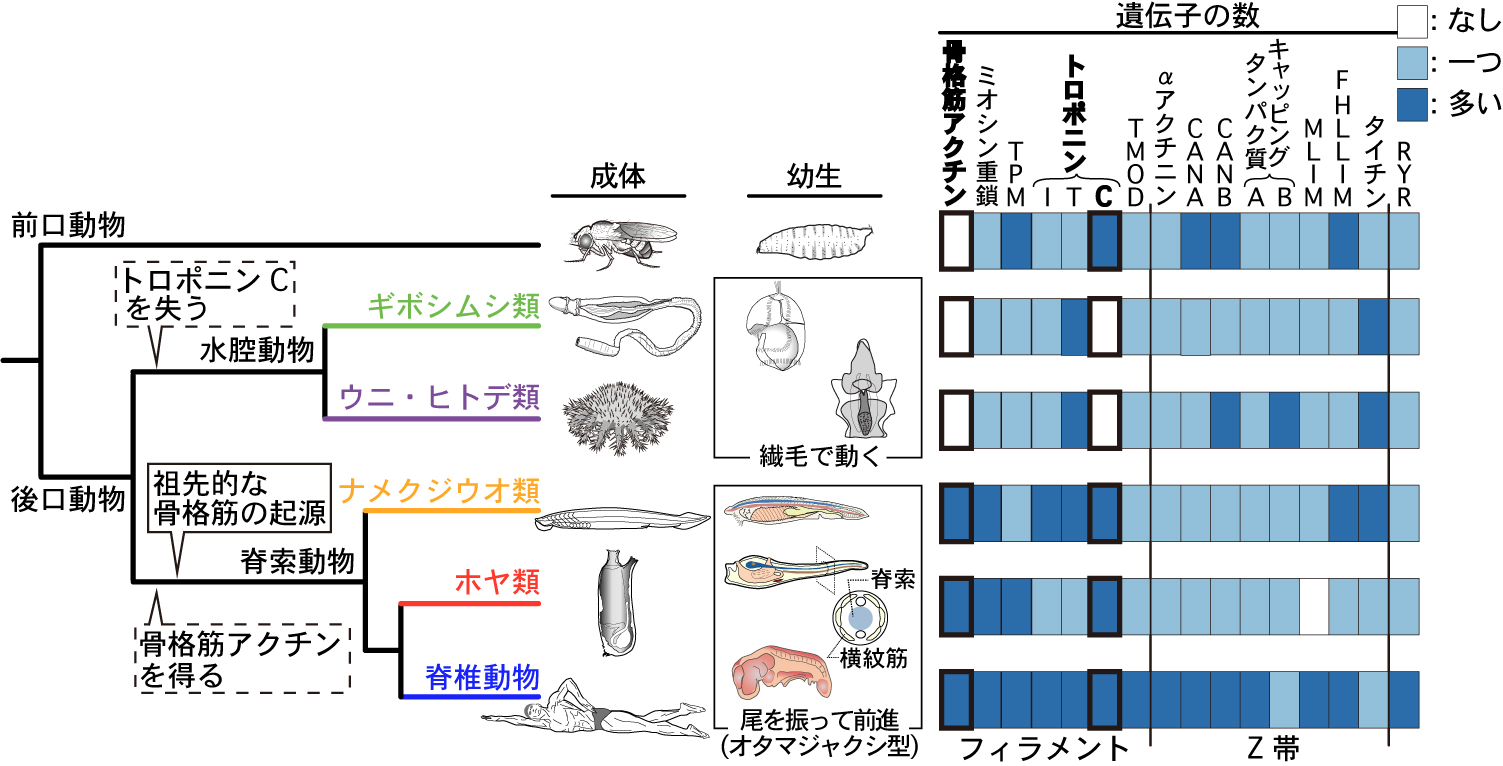 |
| 図2. 後口動物の系統関係と,骨格筋遺伝子と由来の同じ遺伝子の保持パターン. |
骨格筋アクチンとトロポニン C 以外の 14 個のタンパク質遺伝子は,水腔動物にも存在した.水腔動物では,これらの遺伝子がどの筋肉で使われるかわかっていないが,骨格筋に相当する横紋筋が見当たらないのは確かだ.このことは,必要な部品がほぼ揃っていたにも関わらず,水腔動物では骨格筋のような速い筋肉が作られなかったことを示す.一方で遺伝子のコピー数は,水腔動物よりも脊索動物で多く,とくに脊椎動物で急増していた.このことは,脊椎動物の骨格筋は,部品のコピー数を系統内部で独自に増加させて発達したことを示す.さらに別の系統に目を向けると,ナメクジウオ (脊索筋) やホヤ (体側筋) の成体だけにある特殊な筋肉では,祖先的な骨格筋タンパク質から派生したタンパク質が利用されていた (詳細は本文).
以上をまとめると,骨格筋の祖先となった横紋筋は,レールとなった骨格筋アクチンとスイッチとなるトロポニン C を特別な部品として,残りは他の器官の部品を代用して獲得されたことになる.
Deuterostome genomics: Lineage-specifi c protein expansions that enabled chordate muscle evolution. Molecular Biology and Evolution,35:914-924. Web.
 |
|